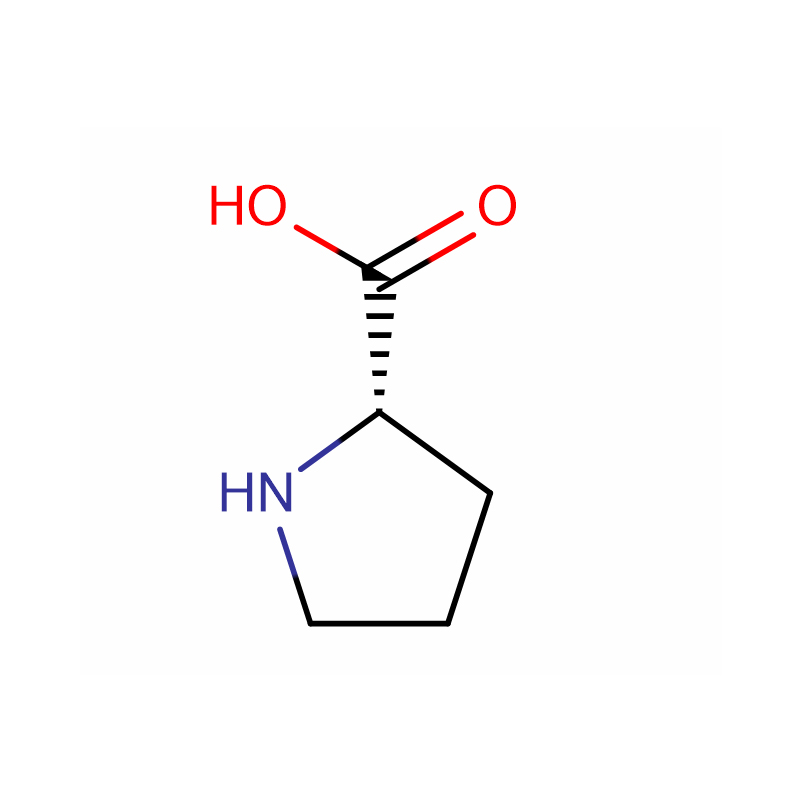
L-Proline Cas: 147-85-3 99% белы парашок
| Нумар па каталогу | XD90293 |
| Назва прадукту | L-пралін |
| CAS | 147-85-3 |
| Малекулярная формула | C5H9NO2 |
| Малекулярная маса | 115,13046 |
| Дэталі захоўвання | Эмбіент |
| Гарманізаваны тарыфны кодэкс | 29339980 |
Спецыфікацыя прадукту
| аналіз | 99% мін |
| Знешні выгляд | Белы парашок |
| Удзельная ратацыя | -84,5 да -86 |
| Цяжкія металы | <15 частак на мільён |
| AS | <1 праміле |
| Ph | 5,9 - 6,9 |
| SO4 | <0,050% |
| Fe | <30 частак на мільён |
| Страты пры сушцы | <0,3% |
| Рэшткі пры гарэнні | <0,10% |
| NH4 | <0,02% |
| Cl | <0,050% |
| Стан рашэння | >98% |
Разуменне метабалізму мікробаў-гаспадара мае важнае значэнне для распрацоўкі і аптымізацыі біякаталітычных працэсаў на аснове цэлай клеткі, паколькі гэта вызначае эфектыўнасць вытворчасці.Гэта асабліва актуальна для акісляльна-аднаўленчага біякаталізу, дзе выкарыстоўваюцца метабалічна актыўныя клеткі з-за эндагеннай здольнасці гаспадара да рэгенерацыі кофактара/косубстрата.Рэкамбінантная кішачная палачка выкарыстоўвалася для звышпрадукцыі пролін-4-гідраксілазы (P4H), дыяксігеназы, якая каталізуе гідраксіляванне вольнага L-праліну ў транс-4-гідраксі-L-пралін з a-кетоглутаратам (a-KG) у якасці косубстрата.У гэтым цельноклеточном біякаталізатары цэнтральны метабалізм вугляроду забяспечвае неабходны косубстрат a-KG, звязваючы біякаталітычныя характарыстыкі P4H непасрэдна з метабалізмам вугляроду і метабалічнай актыўнасцю.Ужываючы інструменты эксперыментальнай і вылічальнай біялогіі, такія як метабалічная інжынерыя і (13)C-метабалічны аналіз патоку ((13)C-MFA), мы даследавалі і колькасна апісалі фізіялагічную, метабалічную і біяэнергетычную рэакцыю цэлаклетачнага біякаталізатара. да мэтанакіраванай біяканверсіі і вызначыў магчымыя метабалічныя вузкія месцы для далейшай рацыянальнай распрацоўкі шляху. Штам E. coli з дэфіцытам дэградацыі праліну быў сканструяваны шляхам выдалення гена putA, які кадуе праліндэгідрагеназу.Біятрансфармацыі цэлай клеткі з дапамогай гэтага мутантнага штаму прывялі не толькі да колькаснага гидроксилированию пролина, але і да падваення хуткасці адукацыі спецыфічнага транс-4-L-гидроксипролина (hyp) у параўнанні з дзікім тыпам.Аналіз патоку вугляроду праз цэнтральны метабалізм мутантнага штаму паказаў, што павышаная патрэба a-KG у актыўнасці P4H не ўзмацняе паток, які стварае a-KG, што паказвае на строга рэгуляваную працу цыкла TCA у вывучаных умовах.У штаме дзікага тыпу сінтэз і каталіз P4H выклікалі зніжэнне выхаду біямасы.Цікава, што штам ΔputA дадаткова кампенсаваў звязаныя з гэтым страты АТФ і НАДН за кошт зніжэння патрэбнасці ў энергіі для падтрымання пры параўнальна нізкіх хуткасцях паглынання глюкозы замест павышэння актыўнасці ТЦА. Выяўлена, што накаўт putA ў рэкамбінантнай E. coli BL21(DE3)(pLysS) быць перспектыўным для прадуктыўнага каталізу P4H не толькі з пункту гледжання выхаду біятрансфармацыі, але і адносна хуткасці біятрансфармацыі і паглынання проліну, а таксама выхаду hyp на крыніцу энергіі.Вынікі паказваюць, што пасля накаўту putA сувязь цыклу TCA з гідраксіляваннем праліну праз косубстрат a-KG становіцца ключавым фактарам, які стрымлівае і мішэнню для далейшага павышэння эфектыўнасці залежных ад a-KG біятрансфармацый.